
Заселение Евразии неоантропами.
Заселение планеты Земля неоантропами разделятся на два разных, но взаимно связанных процесса – миграции носителей определенных генов или языков . Если отдавать предпочтение изучение одному из этих процессов, то результаты будут значительно отличать между собой. В этом можно убедиться, ознакомившись с работой венгерского ученого (Horvát Csaba-Barnabás. 2021) и предлагаемой здесь. Предвидя это, я постарался связать между собой результаты лингвистических, археологических, антропологических и генетических исследований. Однако палеогенетика еще только зарождается как наука, поэтому определенные выводы делать еще рано.
Следы первых людей в Европе обнаружены возле села Вертешселеш в Венгрии и в итальянском городе Изерния, которые датируются 850-700 гг. до Р.Х. (Дейвіс Норман, 2000, 87). Существует точка зрения, согласно которой южные области Восточной Европы впервые были заселены людьми шелльской (аббевильской) культуры около 700 тысяч лет тому назад перед началом миндельского (окского) ледникового периода. Это территория вокруг Черного моря, где было тепло даже в зимние месяцы года (Рыбаков Б.А., 1966, 18). Однако датировка найденных стоянок спорна и не исключено, что они относятся к следующему, ашельскому периоду. В пределах Украины памятники ашельской культуры обнаружены в нескольких местах по берегам Днестра, в частности возле сел Лука-Врублевецкая и Бабин (Толочко П.П., 1994, 14). Данные о ашельских памятниках в других частях Восточной Европы отсутствуют. В дальнейшем произошло, по крайней мере, еще два обледенения и три периода потепления, во время которых люди то отходили на юг, то возвращались на север снова. Что-либо определенное об их роли в этногенезе человека современного типа сказать нельзя.
В Азии первый человек появился значительно позднее, чем в Европе, но, судя по археологическим находкам, в своем проникновении на Восток он достиг Алтая приблизительно 300 тыс. лет назад. Междисциплинарные исследования показали, что пещера Денисова, расположенная на правом берегу реки Ануй в Алтайском крае Российской Федерации, была занята людьми от раннего среднего палеолита до позднего средневековья. Генетический анализ кости пальца ребенка, обнаруженной в пещере, выявил неизвестный тип человека, известный как денисовцы (Berkant E.B. 2017: 499).
Как показали исследования, ДНК фаланги пальца ребенка отличалась от генетической структуры современных людей в два раза большим количеством позиций, чем ДНК неандертальца отличается от нашей. Из этого следует, что денисовцы и неандертальцы следовали разными эволюционными путями дольше, чем неандертальцы и современные люди. Расчеты дали основание предположить, что около миллиона лет назад от Homo erectus в Африке произошли две отдельные линии. Одна из них дала начало неандертальцам и современным людям, другая эволюционировала в денисовцев в Азии (Krause Iohannes mit Trappe Thomas. 2020, 25).
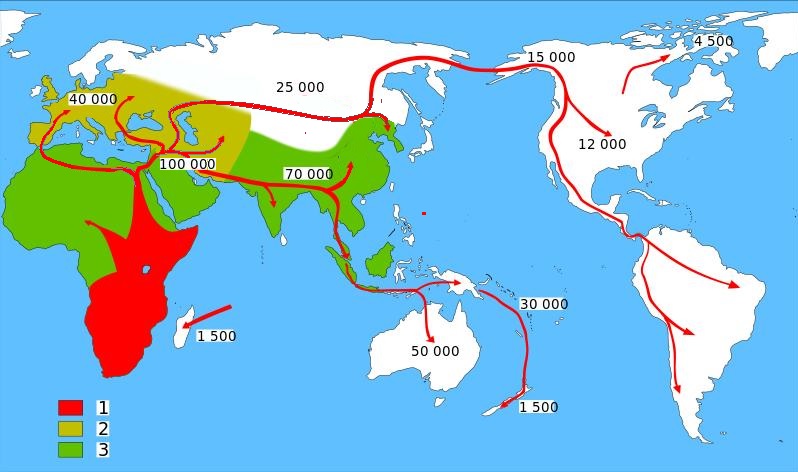
Слева: Распространение homo sapiens.
1. Homo sapiens sapiens.
2. Homo sapiens neanderthalensis.
3. Ранние гоминиды
Человек разумный Homo sapiens, ныне существующая человеческое существо впервые появилась, возможно, 250 тыс. лет назад в Африке.
Генетические исследования показали, что "базовая анатомия Homo sapiens присутствовала там по крайней мере 150 тыс. лет назад" (Stringer Chris. 2007, 15). А 100 тыс. лет назад ранние современные люди были распространенв в Леванте. В это же время там присутствовали неандертальцы:
… потенциальное перекрытие появляющихся современных и неандертальских кладов (покрытий – ВС) было постоянной чертой региона в конце среднего плейстоцена, а также в начале позднего плейстоцена (там же, 16).
Из Леванта в Европу люди стали прибывать более 40 000 лет назад и долгое время сосуществовали с неандертальцами, процесс вымирания которых растянулся на тысячелетия и охватил обширную географическую область (там же, 17). В этом процессе все еще остается для нас много неясного, поэтому мы его касаться не будем, тем более, что он выходит за рамки нашей темы.
По комбинациям антропологических признаков специалисты выделяют среди предков современных людей, заселявших Европу, два (или даже три) типа и связывают с ними территориальные «сгущения» верхнепалеолитических памятников. Одно из них локализуется во Франции по долинам крупных рек, а другое в Центральной Европе на территории современной Чехии и прилегающих областей (Бромлей Ю.В., 1986, 25). По старой, но устоявшейся терминологии их обобщенно называют кроманьонцами. Антропологически кроманьонцы отличались от современных людей разве что более массивным строением тела, но это касается древнейших представителей Homo sapiens. Со временем, в процессе культурного развития (переход на мясную пищу и приготовления ее на огне и т.п.) в строении тела человека происходили определенные изменения, пока оно не приобрело современный вид ( Елинек Ян, 1972, 110).
Для человека того времени характерно ведение присваивающего типа хозяйства с преобладанием охоты и собирательства, но с потеплением климата условия для охоты начали ухудшаться вследствие сокращения площади лесов и наступлением сухой зоны с юга. Кроме того, что потепление заставляло людей двигаться дальше на север и северо-восток, уменьшение продовольственной базы в виде крупных млекопитающих обусловило рост значения рыболовства, особенно в районах многочисленных озер и болот. Люди стали селиться, сначала только летом, а затем и постоянно, вдоль берегов рек и озер, а позже и на берегу Балтийского моря, то есть там, где они могли ловить рыбу (Sahrhase Dietrich, Lungbeck Johannes. 1992, 8). Значительное изменение в структуре присваивающего хозяйствования местного населения имело следствием формирования особых черт в его этнопсихологии, в большой степени определивших его дальнейшее культурно-историческое развитие.
Реки играли и другую важную роль в жизни древнего человека. Направление движения людских потоков в большой степени определялось именно строением гидрографической сети и облегчалось существованием природных условий для прокладки постоянных троп:
Тропы в глубокой древности возникали, прежде всего, вдоль водных потоков вследствие того, что после поселения человек своими непроизвольными усилиями прокладывал пешеходные тропинки чаще всего на ближайшей к водному потоку террасе… Существование иных возможностей прохождения местности по другим линиям из позднейших времен, например, «по верхней трассе», целиком по сухой земле ничего не меняет в способе прокладывания основных трасс. Предпосылки для возникновения пешеходных троп были, прежде всего, физикогеографическими. Социогеографические аспекты возникают вследствие передвижения человека по тропам и образа жизни возле них. Тропы образовывали основной скелет места проживания человека, и не только коммуникативный. Самым важным проявлением сети троп явилось возникновение и продолжительное существование селений (Květ Radan. 2000, 295 )
До начала мезолита общее количество населения Земли составляло только несколько миллионов человек, а в начале неолита (VII тыс. до н.э.) – около 10 миллионов. (Козлов В. И., 1982, 12). Преобладающая часть населения концентрировалась тогда в Южной и Восточной Азии, Африке и Южной Европе, а огромные пространства северной половины Евразии оставались почти полностью безлюдными.
Территорию же Восточной Европы кроманьонцы начали заселять 15 – 12 тыс. лет назад, как только закончился последний период оледенения вследствие резкого потепления и уменьшения континентальности климата. Первоначально это были охотники на больших млекопитающих: мамонта, слона, волосатого носорога. Потепление не было постепенным, а имели место периоды похолоданий из-за новых наступлений ледника из Скандинавии. Однако такие пульсации ледника ослабевали по мере удаленности от Балтии, поэтому степная природно-ландшафтная зона мало изменилась с момента своего формирования в конце третичного периода(Залізняк Л.Л. 1995-1, 3). Существуют классификации климатических периодов, составленные разными исследователями, которые несколько отличаются одна от другой. Ниже представлена таблица периодизации финального палеолита по данным Залізняка(там же)
| 8900 г. до н.э. – 8100 г. до н.э. | Дриас ІІІ |
| 9800 г. до н.э. – 8900 г. до н.э. | Аллерёд |
| 10000 г. до н.э. – 9800 г. до н.э. | Дриас ІІ |
| 10400 г. до н.э. – 10000г. до н.э. | Бёллинг |
| 11300 р. до н.е. – 10400 р. до н.е. | Дріас І |

Слева: Таблица периодизации финального палеолита.
Справа: Дриада восьмилепестковая (лат. Dryas octopetala L, фото из Википедии).
В процессе постепенного отхода ледника на север на его месте образовалась приледниковая тундровая зона, холодные и сухие степи с доминированием в растительности дриады восьмилепестковой, поэтому этот период был назван дриасовим. В фауне доминировал северный олень, который стал важнейшим продуктом питания людей того времени. (Залізняк Л. Л., 1995-1, 3)
Далее на юг развивалась лесная растительность, представленная в основном сосняками и березняками. Одновременно сокращалась роль травянистых и кустарниковых формаций, типичных для позднеледникового времени (Хотинский Н.А., 1977, 57).
Изменение природных условий вызвали, по мнению Шумкина, кризис специализированного хозяйства населения Центральной Европы, состоявшее из охотников и собирателей, которые теперь были вынуждены приспосабливаться к новым экологическим условиям между тундрой и лесом, и это препятствовало быстрому росту производительных сил (Шумкин В. Я, 1990, 10). Кроме того, тут не было таких видов растений и животных, которые бы годились для культивации и доместикации. Все это способствовало перемещению людей в поисках новых охотничьих угодий. На основании анализа археологических находок есть основания говорить, что общее направление движения групп первых охотников в Европе было с запада на восток (Sahrhage Dietrich, Lundbeck Johanes, 1992, 15).
Одновременно начался переход к охоте на мелких животных, который был подготовлен развитием орудий труда, в первую очередь метательного оружия в конце палеолитической эпохи (Формозов А. А., 1959, 68). Человек уже знал лук и стрелы, поэтому мог охотиться не только методом загона на больших млекопитающих, которые в то время же уже начали исчезать, но и на меньших, таких как олени, лоси, медведи, косули, зайцы. Это происходило на финальной стадии культур типа Мадлен (XV – X тыс. до н.э.), центр которой лежал в Юго-западной Франции и в Восточной Испании, но элементы которой можно обнаружить в Германии и Польше (Брей У., Трамп Д., 1990). Очевидно в Северной Германии и Нидерландах развитием мадленской традиции была аренбургская культура, существовавшая на протяжении IX тыс. до н.э.

Идя за северным оленем (см. фото из Википедии слева), который в процессе таяния ледника отходил далее на север, группы охотников приледниковой Европы продвинулись с запада на территорию современной Польши, а с юга или юго-запада – до Белоруссии и Литвы. Здесь развились две поздепалеолитическо-мезолитические культуры – свидерская и несколько восточнее от нее – красносельская, на основе которой позднее развилась песочноровская культура.
Свидерская культура в IX – VIII тыс. до н.э. концентрировалась в районе Литвы, занимая часть Польши, Белорусию (неманская и посожская культурные группы), на юге достигала Карпат, а некоторые ее элементы встречаются даже в Крыму. Восточная граница этой культуры достигала верховьев Днепра и Волги (Римантене Р. К., 1971, 70, 117; Кольцов Л. В., 1977, 119; Мейнандер К. Ф., 1974). Причиной формирования на этих территориях сосредоточия мезолитических культур можно объяснить не только приледниковой зоной, которой придерживался северный олень, но и наличием сырья для изготовления орудий труда и оружия для охоты. Как указывал Калечиц, территория Белоруссии очень богата на легкодоступные залежи кремниевого сырья, особенно в пределах Белорусского Поднепровья (Калечиц Е. Г., 1984, 16).
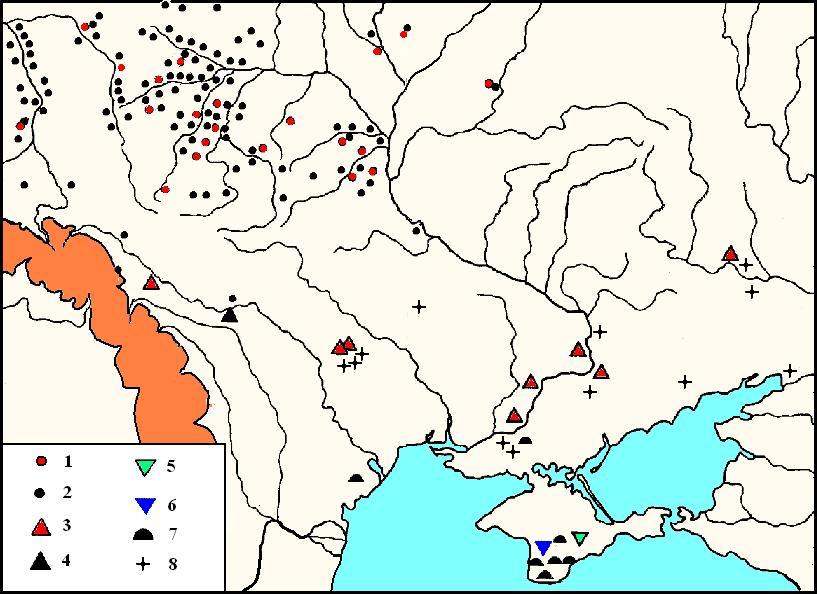
Памятники финального палеолити (ХІ – ІХ тыс. до н.э.
Арта скомпонована на базе данных Зализняка (Залізняк Л.Л. 1995, 4. Рис. 1.)
Цифрами на карте обозначены памятники культур: 1 – красносельськая. 2 – групп свидерской. 3 – осокоровской. 4 – молодовской. 5 – Сюрень. 6 – Вишенне. 7 – Шан-коба. 8 – Восточный Гравет.
Со временем население аренсбургской традиции продвинулось далеко на север Фенноскандинавии (культура комса), а также в Прибалтику и южные районы Финляндии (культуры кунда и суомосярви). При этом существенную роль в формировании последних сыграли носители постсвидерских традиций (Шумкин В. Я, 1990, 11). С появлением этих культур в Европе начинается мезолитический период, который в целом совпадает с переходом в истории Земли от ледниковой эпохи плейстоцена к геологической современности голоцена. Считается, что мезолит продолжался в Восточной Европе с IX до VI тыс. до н.э.. (Археология Украинской ССР).
Постепенно двигаясь на северо-восток, носители свидерских традиций в VII – VI тыс. до Р.Х. достигли бассейна Оки, и, кажется, продвинулись даже до Камы, приняв участие в создании ряда так называемых циркумполярных культур. Эта группа родственных культур в лесной зоне Европы была распространена на севере от зоны, в которой было возможно оседлое земледелие. По словам Чайльда Гордона все без исключения лесные культуры бореальной фазы ведут свое происхождение от верхнепалеолитических культур Восточной и Центральной Европы (Чайлд Гордон, 1952, 33). Уже когда на юге установился неолит, население этой зоны продолжало заниматься охотой и рыболовством. Рыбацкие сетки эти люди плели из лыка, поскольку липа, как и вяз широко мигрируют на север с началом потепления в бореальном периоде, т.е. с VI тыс. до Р.Х. (Хотинский Н.А. , 1977, 59). Позднее они научились от южных соседей гончарному производству и изготовляли яйцевидные сосуды с ямочным или гребенчатым орнаментом (Брей У., Трамп Д., 1990).
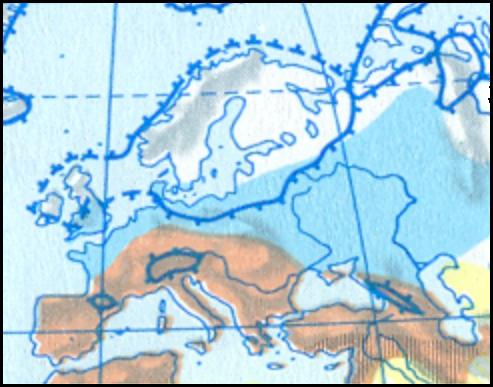
Слева: Заселение Европы после последнего оледенения в 10-9 тыс. до н.э..
Карта взята из "Атласа к истории" [Berthold Lothar (Leiter). 1973. 2.IIІ].
Расселение охотников приледниковой Европы показано синим цветом.
Заселение Восточной Европы в эпоху финального палеолита и раннего мезолита проходило как из Центральной Европы, так и со стороны Балкан и Кавказа в разное время, поэтому определить, какие из культур были автохтонными, а кикие привленесенными, опредить сложно, По этому поводу среди археологов могут быть разные мнения.
В свое время А. А. Формозов выделил на территории Восточной Европы времен мезолита три этнокультурные области – южнорусскую, западнорусскую и центральную (Формозов А. А., 1959, 71). Характерной чертой южных мезолитических стоянок он считал оружие геометрических форм. Такие формы были распространены в Крыму, на Кавказе, в районе Днепровских порогов, на средний Донщине. Западная граница области проходила где-то между Днепром и Днестром, на востоке – между Волгою и Уралом. На юге кавказские стоянки с геометрическими фигурами смыкаются с аналогичными памятниками в Иране. Северная граница проходила через средний Северский Донец и окрестности Полтавы. Для среднерусской области характерны стоянки с наконечниками из ножевидных пластинок. Район распространения – бассейны Оки, верхней Волги, Десны, Белоруссия и Литва. Эти стоянки смыкаются с польскими стоянками свидерской культуры, однако полного тождества здесь нет, поэтому Формозов разделят всю эту область на две группы – "западнорусскую" и окскую (как это допускал в свое время П. П. Ефименко). При этом только у западной группы, которая включает стоянки Литвы и Белоруссии, можно найти много общего с мезолитом Польши (Там же, 75-77). Границы центральнорусской и западнорусских культурных областей остаются неясными. Позднее Формозов отказался от своего трехчленного деления, поскольку геометрические орудия оказались также широко распространенными также в Европе и он признал, что "мезолит, характеризующийся геометрическими орудиями, – явление не столь узкое территориальное" (Формозов А. А., 1977, 59). Действительно, геометрическая форма характерна и для наконечников тарденуазской культуры и эта традиция с территории Франции распространилась через Польшу до Литвы и Белоруссии. Однако внешнее сходство геометрических форм еще не доказывает их общее происхождение. Они могли быть занесены в Восточную Европу как с запада, так и со стороны Кавказа и Балкан, откуда миграция носителей иной этнокультурной традиции, отличной от тарденуазской по другим признакам, должна была иметь место.
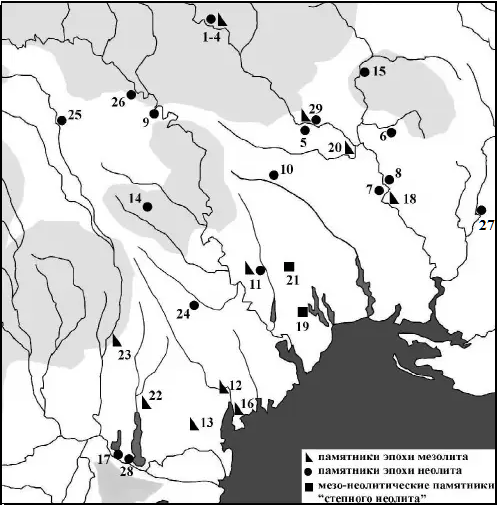
Исследование одесских археологов дают основание утверждать, что носители культур мезолита заселяли Восточную Европу также через Балканы.
Слева: Карта памятников мезолита и неолита Северо-Западного Причерноморья
(З ДКСЗП. 2013, 14).
Условные обозгачения: 1 – скибенецкий узел памятников; 2- соколецкий узел памятников; 3- Заньковцы ІІ; 4 – Печера; 5 – Саврань; 6 – Миколина Брояка; 7 – Гард III; 8 – Пугач II; 9 – сороцкий узел памятников; 10 – Майнова Балка; 11 – Гиржево; 12 – Белолесье; 13 – Мирное; 14 – Сакаровка; 15 – Добрянка 3; 16 – Траповка; 17 – Орловка; 18 – Арбузова Балка; 19 – Карпово; 20 – Конецполь; 21 – Катаржино I; 22 – Зализничное; 23 – Саратены; 24 – Дэнчень; Русештий-Ной; 25 – Бедражий Векь; 26 – Тэтэрэука-Ноуэ XV; 27 – Новорозановка; 28 – Новосельское; 29 – Мельничная Круча.
Геометрические формы орудий, которые были найдены на этих памятниках, говорят, что такие формы могли развиваться в разных местах при использовании подобного метода их изготовления:
Развитой поздний мезолит появляется в Северо-Западном Причерноморье ранее его европейских аналогов (кастельновьена и тарденуаза) – еще в бореальном периоде голоцена. Северо-Западное Причерноморье может рассматриваться как один из центров распространения техники получения правильных призматических пластинок (отжимом и/или через посредник) и симметричных микро-литических наконечников в форме трапеций (Киосак Д.В., Пиструил И.В. 2013, 23).
Таким образом, надежных данных для выделения каких либо этнокультурных областей эпохи мезолита в Восточной Европе археология пока не имеет.

После окончательного таяния ледника в Скандинавии начинается атлантический период, который считается климатическим оптимумом голоцена. На севере тундровая зона сокращается за счет наступления тайги. В Европе распространяются широколиственные леса, даже в степной зоне по долинам рек. Одновременно меняется фауна – исчезают стада северного оленя, на смену которому приходят нестадные копытные – лось, тур, благородный олень (на фото из Википедии справа), кабан, косуля.
Радикальные изменения в природно-ландшафтных обстоятельствах имели следствием и тотальные изменения в этнокультурной ситуации. Ни одна из позднепалеолитических культур Украины не нашела продолжения в раннемезолитических, которых археологи на основании классификации микролитов выделяют пять: Шпан-Коба, Кукрек, Зимовники, Песочный ров и Кудлаевка (Залізняк Л.Л. 1995-2, 4-6)
Памятники культуры Шпан-Коба сосредоточены в горной части Крыма, где также имеются памятники культуры Кукрек, но она больше распространена в причерноморских степях. Эта культура не имеет аналогов в мезолите Европы, если речь вести о кремневых изделиях. Основой экономики Кукрек была охота на копытных животных, но есть находки дающие основания говорить о начале развития среди кукрекского населения животноводства. Влияние этой культуры распространялось в Приазовье и Полесья. Она считается автохтонной (там же, 6-10)
Культура Песочный ров была распространена в центральной части Восточной Европы и имеет несколько вариантов. Она считается продолжением аренсбургской традиции. Некоторые общие черты с песочноровской имеет культура Зимовщики, памятники которой известны в Правобережной лесостепи. Аренсбургские черты, но другого типа, имеет культура Кудлаевка, распространенная на Полесье (там же, 10, 12, 13). Очевидно носители этих трех культур мигрировали в Восточную Европу из Центральной.
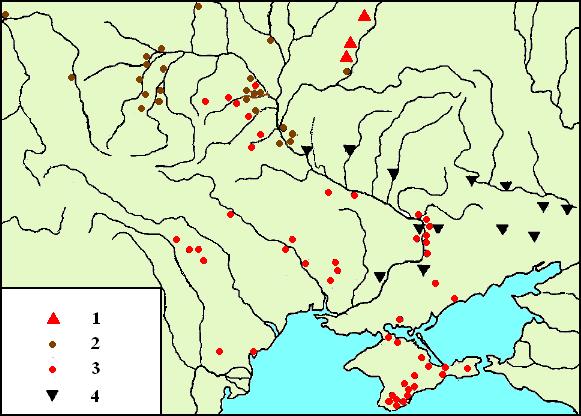
Справа: Памятники раннего мезолита Украины VIII-VII тыс. до н.э.
Оригинал карты (там же, 6. Рис. 1) тонирован цветом автором.
Условные обозначения:
1. – песочноровска культура.
2. кудлаевская культура
3.кукрекская культура.
4.зимниковская культура.
В подавляющим большинстве случаев языки всех эти людей, о которых шла речь, в целом не сохранились. Их относят к палеоевропейским и палеоазиатским. О некоторых народах, говоривших на них, имются скупые исторические сведения, в Европе это были лигуры, турдетаны, возможно, иберы и другие. Ученые пытаются реконструированть языки этих народов при помощи исследований сохранившейся топонимии непонятного происхождения. Кроме топонимии, может помочь языковой субстрат, который должен существовать в современных языках, который можно найти в процессе их этимологизации. При восстановлении єтих языков следует исходить из представления о первых звуках человеческой речи (см. Происхождение человеческой речи). В Азии палеоазиатские языки существовали до новейшего времен, а один из них, кетский, сохранилия до сих пор. Он относится к енисейским языкам, на которыъ гоаорили еще исчезнувашие арины, ассаны, котты. Антропологичнские черты древнейшего населения Евразии сохранились в генетике, а представление об их культуре дают археологические находки.
Данные о заселении Евразии ближайшими предками современных людей по языку были получены а процессе изучения родственных отношений ностратических и сино-тибетских языков при помощи графоаналитического метода. Подобие их графических моделей позволило предположить, что формирование праязыков, давших начало этим большим макросемьям происходило в одном и том же месте. Поскольку ранее было определено, что ностратические праязыки сформировались во времена мезолита на территории Закавказья и Армянского нагорья в районе трех озер Ван, Севан и Урмия, было принято, что те же места заселяли сино-тибетцы. Формирование одельных языков происходило в одних и тех же этноформирующих ареалах вокруг горы Арарат.
В поисках убежища в холодную пору года население Леванта осваивал пещерные жилища на Кавказе значительно раньше, чем соседние районах Ближнего Востока. Кавказ занимает одно их первых мест (а возможно и первое) по количеству стоянок древнекаменного человека (Любин В.П. 1998, 49). Интенсивное заселение этой горной страны сино-тибетцами происходило в раннем палеолите и далее эту традицию продолжалили ностатические народы (см. карту ниже).
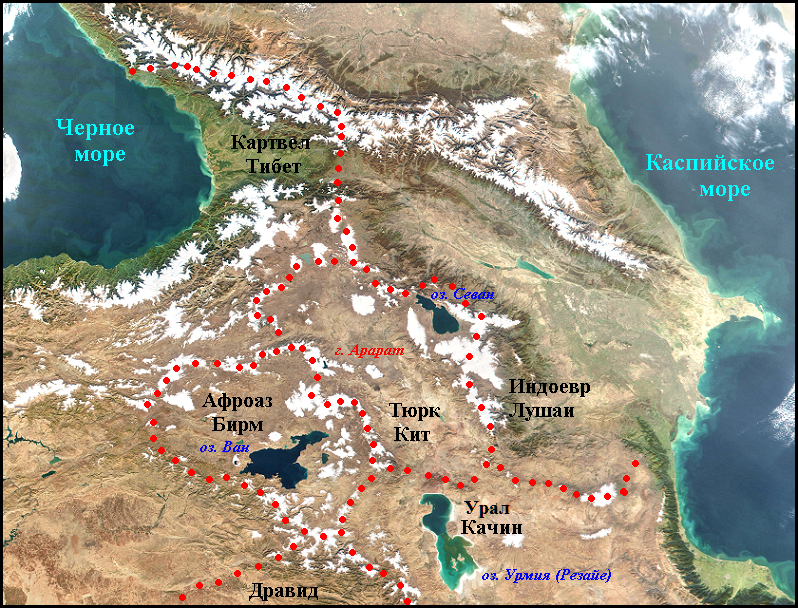
Этноформирующие ареали в район трех озер
Ностратические првязыки: Афраз. – Афразийский, Драв. – дравидийский, Индоевр. – индоевропейский, Картвел. – картвельский, Урал – уральский, Тюрк. – тюркский.
Сино-тибетские языки: Бирм. – бирманский, Кит. – китайский, Тибет- тибетский.
Представляется невероятны, что при переднеазиатском происхождении принадлежащих к желтой расе сино-тибетцев эта раса могла соформирваться где-то вблизи их современных мест расселения под влияним местных природных условий и дадее оаспространиться га огромном пространстве в совершенно иной окружающей среде. Так можно думать следуя логике, но ученый мир склоняется к такой мысли в ходе генетических исследований:
… многие «древние» маркеры ДНК, взятые за пределами Африки и привлеченные для аргументации в пользу генетического происхождения от внеафриканских архаичных видов.., в конечном итоге могут оказаться перенесенными из Африки в ходе современных человеческих расселений, за которыми последовали последующие изменения между Африкой и внешним миром большой частоты (Stringer Chris. 2007, 17).
Продолжая думать логически, следует иметь в вмду, что от специфических монголоидных признаков – узкого разреза глаз и эпикантуса потомкам избавиться трудно. Поскольку при переселении народов какая-то их часть всегда остается на прежних местах обитания, то ассимилированные впоследствии кавказодидами предки сино-тибетцев все-таки должны были иметь некоторые монголоидные признаки. И, как оказалось, свидетельства этому есть. Некоторые исследователи обращали внимание узко посаженные глаза фигур на ранних шумерских рельефах. Для примера см. рис. ниже.

Слева: Статуя знатного шумера. Лагаш. Около 2500 г. до н.э..
Статуя знатного шумера, действительно, демонстрирует несколько суженные глаза мужчины. Кроме того, у фигуры можно предполагать также наличие эпикантуса, складки верхнего века. Это могут быть остаточные признаки метисации кавказоидов с монголоидами. То, что эти признаки выражены довольно слабо, можно объяснить тем, что монголоиды оставили Шумер несколько тысячелетий тому назад.
Является общепризнанным, что шумеры не были семитами, хотя их происхождение принадлежит к числу неразрешенных загадок истории. Шумерский язык имеет так же мало общего с семитскими, как и другими основными известными языками (Krämer Walter, 1971, 67). Идея западноазиатского происхождения сино-тибетцев в разных вариантах не нова и основывается как на определенном сходстве китайских и ранних аккадских иероглифов, так и на обнаруженных лексических соответствиях между китайским и шумерским языками. E.T. Уильямс, как и другие ученые, считал, что у них были некоторые туранские черты, и поэтому пришел к такому выводу:
Таким образом, мы имеем факты в пользу того, что различные китайские племена появляются там, где сейчас находится сейчас Китай, прийдя из какой-то области на северо-западе этой страны, и что шумеры появляются в долине Евфрата, прийдя из какого-то места на северо-востоке Вавилонии; что шумеры были очевидно туранской расы, и что их язык и их письмо поразительно похоже на древнее китайское и что значительные изменения в климате Центральной Азии гнали в разные периоды немалое число жителей, мигрировавших в различных направлениях. В целом не кажется невероятным то, что предки китайцев предки и предки шумеров, возможно, были родственны между собой и, возможно, мигрировали из соседних регионов, китайцы на восток, а шумеры на запад (Williams E.T., 1918: 211).
Время пребывания китайско-тибетского населения на прародине следует отнести к верхнему палеолиту, потому что позже это место было заселено носителями ностратических языков, которые прибыли сюда с запада, оттесняя коренных жителей на восток и ассимилируя или уничтожая их остатки. Сино-тибетцы переселились в Центральную Азию, где они стали создатели местных мезолитических культур. К нынешним местам своего обитания они прибыли уже в неолите или принесли его с собой, так как ни в Китае, ни Бирме надежные следы мезолита не были найдены. Неолитическая культура Яншао в среднем течении реки Хуанхе существовала в V—II тыс. до н. э.

Справа:
Статуя бородатого жреца из Мохенджо-Даро.
Очевидно, люди монголоидной расы оставались в Центральной Азии до прихода сюда дравидийских народов. Наличие монголоидных признаков у жреца из Мохенджо-Даро может подтверждать это предположение
Типологическая неоднородность людей разных рас в позднем палеолите была меньшей, чем теперь. (Щокін Георгій, 2002, 77). Иначе говоря, разница между прототипами людей европеоидной и монголоидной рас была незначительна и проживали они в одинаковых природных условиях Передней Азии. После того, как монголоиды мигрировали по широким просторам Азии, их первоначальные фенотипические черты были развиты различным образом под влиянием природных условий разных мест обитания. В результате их отличие от людей белой расы возросло, но одновременно развились два отличных расовых типа, давших начало как современным монголоидам, так и американоидам.
К желтой расе относятся также индейцы Америки, народы монгольской и тунгусо-маньчжурской языковых групп. При решении вопроса о происхождении этой расы следует обратить внимание на то, что у индейцев Америки, эпикантуса нет, а айны, древнее население Японии, Сахалина и Курильских островов, вообще обладают некоторыми явно выраженными европеоидными признаками. Полный набор монголоидных черт можно найти у населения Юго-Восточной Азии, народов монгольской и тунгусо-маньчжурской групп. Языки последних не содержат четких признаков родства с сино-тибетскими, т.е. формировались очень далеко от поселений сино-тибетцев, поэтому можно предполагать, что люди желтой расы заселяли огромное пространство в Восточной Европе и Азии в то время, когда речь человека проходила только начальный этап своего становления.
По данным антропологических исследований верхнепалеолитических поселений в разных регионах Европы проживали представители всех основних расових групп. Результаты анализа многочисленной эпипалеолитической находки черепов в Баварии дают такие результаты: нордический элемент – 14.4%, лапоноидный – 54.2%, средиземноморский – 31.4% (Czekanowski Jan, 1957, 30). Монголоидные черты отличают население ранненеолитической елшанской культуры, распространенной в междуречье нижней Волги и Урала. По крайней мере некоторые из черепов, найденные в немногочисленных погребениях этой культуры, могут быть отнесены к лапоноидному типу.

Слева: Три саамские женщины. 1890 годы. Фото из Википедии
Поскольку лапоноидная раса является промежуточной между монголоидной и европеоидной, наличие лапоноидного элемента в эпипалеолитических находках черепов в Восточной Европе может свидетельствовать, что в верхнем палеолите здесь происходила метисация антропологически отличных этносов, которая в конечном итоге привела к особому многообразию расовых типов, которые теперь обобщенно относятся к желтой расы.
Генетика дает предварительное объяснение этой метисации. В период неолита Европу сформировали два генетических блока: ДНК давних охотников и собирателей и ДНК земледельцев, мигрировавших из Анатолии. Они сохранились до сегодняю. Однако имеется есть еще один генетический столп, ярко выраженный на севере и востоке Европы, но, по крайней мере, отчетливо заметен на остальной части континента. Еще в большей степени он сохранилися у коренных жителей Америки. Считается, что Америка заселялась через Азию, поэтому генетически она должна быть ближе к Америке, чем Европа, но на самом деле наоборот: генетически европейцы более родственны коренным жителям Америки, чем люди Восточной и Южной Азии (Krause Iohannes mit Trappe Thomas. 2020, 114). Генетические исследования лапоноидов могут объяснить это противоречие.
Расселение людей в те древние времена обусловливалось локальным давлением населения либо за счет его увеличения внутри общины, либо за счет его увеличения среди соседних племен. (Козлов В.И., 1982, 17). Охотники двигались группами численностью около 25 человек. Понятно, что такой коллектив не может быть племенем. Не совпадает он и с отцовским родом. Как указывает Арутюнов, "племя, состоящее из таких коллективов, каждый из которых нуждается при самых благоприятных условиях не менее, чем в 400-500 кв. м. охотничьей территории, а чаще всего территории в 2-3 раза большей, не может быть ни спаянным, ни многочисленным" (Арутюнов С.А. 1982, 65). Таким образом, расселение первобытных людей на широких просторах Восточной Европы само по себе могло больше задерживать их прогресс, чем способствовать ему, поскольку, попадая в результате дальних миграций в незнакомую природную среду, люди должны были тратить время и силы на приспособление к новым условиям существования (Козлов В.И., 1982, 14).
Нарисованная выше картина заселения первичного заселения Восточной Европы является гипотетической и значительно упрощенной. Все эти вопросы далеко не разрешены, поэтому целью этой главы было показать возможные варианты первичных этногенетических процессов до того момента времени, когда о них можно говорить более уверенно. Однако в соответствии с темой этой работы нас будет более интересовать вопрос, могли ли первые собиратели-охотники создавать стойкие этнические группировки. При его рассмотрении будем иметь в виду такие формулировки:
Этническая общность обычно определяется как совокупность людей, имеющих общую культуру, говорящих, как правило, на одном языке и осознающих как свою общность, так и отличие от людей, принадлежащих к другим таким же общностям (Семенов Ю.И., 1986, 73).
Этнос является устойчивой, исторически сложившейся общностью людей, характеризующейся такими признаками-свойствами, как общность территории, языка, экономических связей, культурного уклада и этнического самосознания, выражающееся прежде всего в сознании действительной или мнимой общности происхождения (Королюк В.Д., 1985, 134).
Указанные выше признаки-свойства составляют устойчивую структуру, которая позволяет этносу сохраняться на протяжении длительного времени в разных исторических периодах даже после потери одного из признаков при условии сохранения самосознания. Таким образом, мы приходим к вопросу о роли наследственности в процессе формирования этноса. Сформированные под влиянием окружающей среды определенные черты физического строения и характера передаются от поколения к поколению разными механизмами:
Генетика поведения изучает основы поведения и все, что с ним связано, – психические заболевания, склонность к разводу, политические предпочтения и даже чувство удовлетворенностью жизнью. Эволюционная психология ищет механизмы, посредством которых эти признаки переходят от поколения к поколению. Оба подхода предполагают, что в формировании поведения, мыслей и эмоций участвует природа и воспитание, но в отличии от практики двадцатого века нынче природе отдается предпочтение (Чиксентмихайи Михай. 2008. 89) .
Генетика проявляется на индивидуальном уровне, а эволюционная психология является социальным механизмом, но дейстувают они взаимосвязано. Во времена палеолита и мезолита среди людей господствовал промискуитет, но эволюционная психологиет имела следствием развитие эндогамии, которая привела к созданию определенных этносов, в которых доминируют определенные гены. Принимая во внимание все известные нам факты, следует признать, что первые поселенцы в Восточной Европе стояли на таком низком уровне общественного развития, что не может быть и речи о существовании каких-либо этнических единиц. Однако эти люди образовали антропологическую основу этногенетических процессов, которые начались в более позднюю эпоху.
